Ginkgo biloba
This remarkable order of great antiquity is represented in the present age by a sole survivor ginkgo biloba, which by many paleobotanists is regarded as a “living fossil”. It occurred as far back as Triassic period of the Mesozoic age (about 200,000,000 years ago). The order as a whole consists of 16 genera and many species, all of which except one are extinct. It came into existence during the Permian and achieved worldwide distribution and luxuriance during the existence during the Triassic and Jurassic periods of Mesozoic age. It gradually thinned out and started fading out of existence during the Cretaceous and onwards into the Cenozoic. The sole living representative, G. biloba, owes its existence and has spread in the modern period due to the interest man has taken in this plant. It has been protected., worshipped and cultivated by man. Still, it grows in a wild state in a small and inaccessible area in Southern China (Li, 1956). It has also been reported as a native plant in the province of Chekiang in eastern China. This plant has vegetative and anatomical characters that are purely coniferophyte, but in many reproductive features it resembles the modern cycads; it also resembles the extinct Cordaitales and even Cycadeoids (see Delevoryas, 1972′) in some features. This synthesis of characteristics from various extant and extinct groups in one living plant certainly makes it a treasure for the botanists and stamps it as a “living fossil”. Foster and Gifford (1959) state, “such similarities appear as the result of the retention, by both Cycads and Ginkgo, of certain very ancient patterns of reproduction which probably were shared by many of the Paleozoic and Mesozoic gymnosperms. If these observations are correct, then it follows that the seed must have evolved the ultimate common ancestor of the cycads and Ginkgo, for the common reproductive features of the two groups, are those associated with the seed habit itself.
Characteristic features of Ginkgo biloba
The living Ginkgo biloba reveals the following characteristics :
- The tall trees give a stately appearance and have an excurrent habit.
- The roots penetrate deep into the substratum and constitute a top root system.
- The leaves are fan shaped and deciduous.
- The venation is open dichotomous.
- The leaves arise singly along the terminal branches and bear in their axils buds that grow ‘into characteristic short shoots, which also bear a dense cluster of leaves at their apices.
- The plant is dioecious and slow growing.
- The wood is pycnoxylic.
- The catkin-like inflorescences that bear micro sporangiophores with two to twelve pendent microsporangia arise in the axils of leaves on dwarf shoots.
- The ovules arise in groups from the apices of axillary branches.
- Presence of an endosperm tent pole is characteristic of this living plant and brings it akin to the cordaitales.
- The mature seeds are large, fleshy and about the size of small apricot. The integument is composed of an outer orange colored fleshy portion and an inner hard and stony layer. The fleshy coat is rich in butyric acid and when crushed emits an odour that is unpleasant. The seeds are produced in large numbers and tend to hang on for a month or two after the leaves are shed. A female tree in this late fall attire gives a beautiful look.
- The spermatozoids are motile and bear a spiral band of cilia
Habit and Habitat
Ginkgo biloba is tall tree that may grow up to 100 feet tall with excurrent habit but may achieve irregular form in older trees. The young plants grow erect and straight for the first twenty to thirty years with a strangely spindly growth habit. The lower branches gradually begin to spread up, a few of them becoming quite massive in the course of a century. The leaves are deciduous. This slow growing tree is considered to be one of the best shade trees for future generations. It can grow along the hill slopes or even in the plains.
External characters:
Stem:
The stem is erect, tall, slow growing and branched. The branching is lateral. There are two types of branches:
1. Long Shoots: They arise from the main stem and grow rapidly and may again bear long shoots and dwarf shoots. The long shoots bear green leaves that arise singly from a node and are spirally arranged with a bud in their axils. They bear deeply lobed foliage leaves.
2. Dwarf Shoots or Spur Shoats: They develop from the axils of foliage leaves on the long shoot and grow very slowly producing each year a small crown of foliage leaves and the reproductive structures. The foliage leaves borne on them are not deeply lobed and are more or less entire. They simulate the pinnae of the fern Adiantum. The dwarf shoots may persist as such for years. They differ from the long shoots in their short length, in anatomical details and also in being structures that bear reproductive organs.
The long shoots and the dwarf shoots are interconvertible. The dwarf shoots have been observed to grow into long shoots if the apical bud of the subtending long shoot is removed, but cases are not rare when the dwarf shoots grow as long shoots without removing the apices of adjoining long shoots. Similarly long shoots have been observed to grow as dwarf shoots for a year or two and then revert to long shoot growth phase. Sometimes after an active growth of one or two years, the long shoot may permanently enter dwarf shoot phase. These observations suggest that there is no fundamental difference between the shoot apical meristem of both kinds of branches. The organization of their apical meristem is also similar. Cunckel and Wetmore (1946) describe that, all the buds on a tree grow into short shoots every spring and some of them change into long shoots during the course of the season. It has also been suggested that differences in the growth pattern of the two shoot types may be due to quantities of auxins produced in the apical meristems (Bierhorst, 1971). In the early spring, the apices of both kinds of shoots produce similar quantities of auxins and as growth proceeds, the auxin production increases. At a defied stage, the auxin production is comparatively more in the long shoots which grow more and thus increase in length. The fact that reproductive organs are borne only on dwarf shoots seems to be related to the production of some specific substances.
Root –
The tap roots are extensively branched and grow quite deep into the soil. The root caps and root hair are present.
Leaves
The foliage leaves of Ginkgo biloba have a characteristic shape, which distinguish the tree from all other gymnosperms. They are simple, large and fan-shaped with expanded apex and narrow base. They are petiolate and are highly variable. When the leaf emerges from the bud, the two lobes of the leaf are folded in towards each other, but it becomes flattened out later. In the early stages, the leaf is pale yellow-green in color, but it becomes typical dark green about half way through its development. During this color change, it becomes more fan-shaped and both the lobes of the leaf increase in area. They are deeply bilobed on the long shoots and on the seedling and may also show secondary lobing. On the dwarf shoots, they are not deeply lobed and may be almost entire or sinuate. They have an open dichotomous venation, there being no interconnecting cross veins. The leaves usually fall during the autumn and turn beautiful golden-yellow before leaf fall. Their venation is straight like those of other conifers. Another feature of the leaf, besides leaf shape, that distinguishes the Ginkgoales from other conifers is the shape of epidermal cells. The epidermal cells between the veins are polygonal in outline whereas over the veins they are rectangular in shape. The walls of epidermal cells are wavy. The stomata are restricted to the lower epidermis and are surrounded by 4-6 subsidiary cells that form a circle around it and have characteristic finger-like projections overarching the guard cells. The stomata occur in between the veins and are arranged in broad bands or ill-defined rows. Kanis and Karstens (1963) have described the presence of stomata on the upper surface of leaves in the long shoots of male plants. Two vascular bundles enter at the base of leaf and undergo repeated dichotomy.
Foster (1 938) studied the shoot apices of Ginkgo biloba. A shoot apex consists of a few superficial apical initials. Below them is the sub-apical zone consisting of a group of central mother cells. Around the proximal part of this zone is a zone of rib meristem. There is no distinct tunica region because the superficial cells usually undergo a periclinal division. The rate of division in sub-meristem cells determine whether the branch will be a dwarf shoot or a long shoot.
Anatomical characters
The anatomical details of the species also present certain interesting features that are discussed below:
Stem
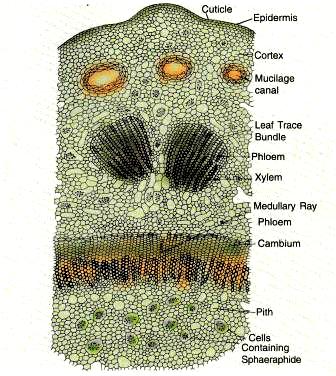
A cross section of the young stem presents a roughly circular and wavy outline. There are some variations in the internal structure of a dwarf shoot and the long shoot.
The outermost layer of the stem is the epidermis which is quite distinct, in the young stems that have not undergone secondary thickening. It is made up of a single layer of brick-shaped cells and is covered with a thick cuticle. In older stems, the epidermis is replaced by periderm which originates in the cortex.
Next to the epidermis is the cortex, which is quite extensive in the dwarf shoots and comparatively narrow in the long shoots. The outer cortical cells contain crystals of calcium oxalate and tannin filled cells. Large resin canals are found scattered in the inner cortical layers. There are no distinct endodermis and pericycle.
The very young stem consists of a number of discrete primary vascular bundles arranged in a ring. These bundles are conjoint, collateral, open and endarch. They run longitudinally throughout the stem and branch sympodially to give off leaf traces. Each leaf receives two leaf traces that originate from two different primary cauline strands (Gunkel and Wetmore, 1946). These leaf traces arise by tangential division of the cauline strand. Soon secondary growth starts and the vascular ring becomes complete and appears to be siphonostelic. The medullary rays are uniseriate and 1-5 cells high in dwarf shoots and 1-15 cells in long shoots. The stelar system in Ginkgo as in all other living gymnosperms, is a eustele and this condition is very clear in young stems. The protoxylem has spiral thickenings whereas the tracheid’s of metaxylem have bordered pits on their radial walls. The phloem is distinct and consists of sieve tubes and little phloem parenchyma. The protophloem gets crushed.
In the centre of the stem is the pith which is large and extensive in dwarf shoots and narrow in long shoots. The pith is traversed by mucilage canals and its parenchymatous cells also contain crystals of calcium oxalate.
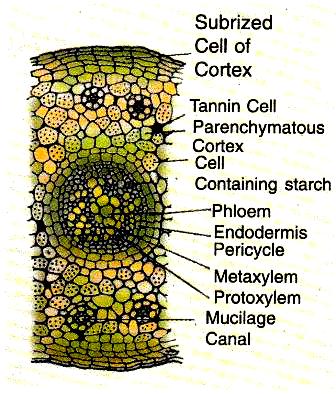
Root
The young root has an extensive cortex made up of thin-walled cells that contain tannin filled cells, mucilage cavities and calcium oxalate crystals.
A distinct endodermis is evident in the young roots. Next to it is the single layered pericycle. This distinction is lost in mature roots.
The young roots are diarch or triarch i.e., there may be two or three xylem strands alternating with the same number of phloem stands. The xylem is exarch. Triarch roots are found in young seedling with three cotyledons.
As a result of secondary growth, the root shows indistinct annual rings. The phellogen develops in the outer cortical layer and from the outer corky layers of root.
Leaf
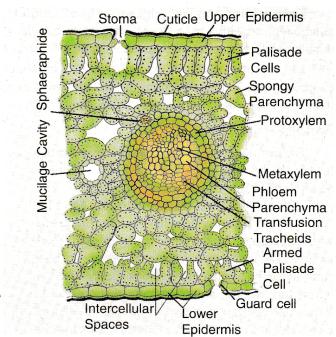
A vertical section through the leaf reveals the upper and lower epidermal layers that ore made up of thin-walled cells. A distinct cuticle covers the epidermal layers. The cells of the epidermis are polygonal between the veins whereas over the veins they are rectangular in outline. The stomata are restricted to the lower epidermis and are haplocheilic. Kanis and Karstens (1963) reported some stomata in the upper epidermis of leaves of long shoots in male plants. The guard cells of the stomata are surrounded by a ring of 4-6 or 7 accessory cells whose surface projects into finger-like out-growths that overarch and guard cells. The cells of the lower epidermis have slightly wavy outer walls. The stomata are arranged in broad but indistinct bands between the veins.
Between the two epidermal layers is the mesophyll tissue that shows no distinction into palisade and spongy parenchyma except in old and mature leaves of some long shoots. The mesophyll cells contain abundant chloroplasts and are loosely arranged so as to enclose small air spaces. Mucilage ducts also traverse the mesophyll. Tannin-filled and crystal-containing, cells also occur in this region.
There is a distinct sclerenchyma Tous bundle sheath (only in mature leaves) around each vascular bundle representing a vein. Sclerenchyma Tous bundle sheath is not developed in young leaves. The bundle sheath extensions are parenchymatous. The leaf otherwise lacks sclerenchyma Tou’s tissue.
The vascular bundle of the vein has a weakly mesarch xylem. Majority of the bundles show endarch xylem. In the cotyledons the vascular bundles are mesarch at the base but become exarch at the distal ends.
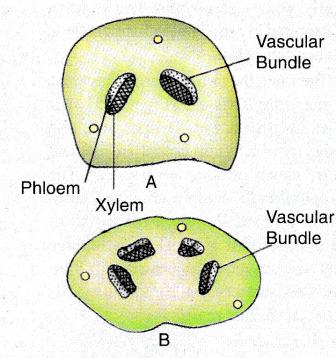
Petiole
The leaf traces are endarch when they enter the petiole but become weakly mesarch or remain endarch during their course through the lamina. The petiole has two endarch vascular bundles. The protoxylem has spirally thickened walls as compared to the pitted tracheids of the metaxylem. The vascular bundles in the petiole are also surrounded by a sclerenchymatous sheath. The xylem is traversed by uniseriate rays, that are one to three cells in longitudinal extent in the bundles of long shoot and its leaf traces, and one to fifteen cells high in the bundles of dwarf shoot and its leaf traces. The medullary rays of xylem are continuous with those of the phloem. The leaf trace bundles have a distinct cambium layer.
Secondary growth in stem
It takes place by the activity of a single ring of cambium that remains active throughout the life of the plant. The wood is pycnoxylic as well as monoxylic. The cambium develops into a complete ring and cuts off secondary phloem elements towards the outer side and secondary xylem towards the pith. It is made up of fusiform initials as well as ray initials. The former give rise to the vascular elements whereas the latter give rise to uniseriate rays that are 1-5 cells high in long shoots and up to 15 cells high in dwarf shoots. The secondary phloem consists of sieve elements and scanty parenchyma. Phloem fibers and companion cells are absent. The secondary xylem consists of tracheid’s and exhibits weekly developed annual rings. The tracheid’s have moderately thick walls and the trochoidal overlap is inexpensive and most of the tracheid’s end the same level. This is responsible for making the wood brittle and of no economic value. The tracheid’s have circular bordered pits arranged in one or two rows and are restricted to the radial walls. The pitting may be alternate or even opposite. The pits have a distinct torus and bars of Sanio are also present. The bars of Sanio are thickened lines that separate the pits and are present in the lumen of the tracheids. They do not cross the lumen or cavity of the trochoidal cell. There also occur trabeculae of Sanio that cross the trochoidal lumen and can, therefore be easily distinguishable from the bras. The tracheids are of moderate length as compared to very long coniferous tracheids.
Cork cambium appeals in the outer cortex and forms a well-developed periderm that replaces the epidermis. This cambium also forms secondary cortex towards the inner side. mucilage canals are absent from the secondary cortex, but tannin cells and calcium oxalate crystals are present in smaller amounts.
Reproduction:
3. Reproductive structures- Ginkgo biloba is monosporangiate and strictly dioecious. Male and female strobili occur on male and female plants respectively.
(a) Male i.e. staminate strobilus-The male strobilus is formed in loose catkin-like cluster at the apex of on male plant. They arise in the axils of foliage leaves or inner bud scales. It consists of a central axis upon which microsporophyll’s are arranged spirally.
A mature microsporophyll has a slender stalk which is turned afterwards into a knob-like or hump-like enlargement, beneath one side of which usually two pendent microsporangia are borne, the hump or knob-like enlargement of the stalk of the microsporophyll is considered by some authors as a sterile third microsporangium. Sometimes 3 to 4 microsporangia have been obtained. Development of the microsporangium is of eusporangiate type. From the archesporial cell, primary wall cells and the sporogenous cells are formed. The sporogenous cell i.e. sporogenous tissue by repeated divisions form the microspore mother cells. microspores i.e. pollen grains are formed from the microspore mother cells by reduction division. Microsporangia dehisce through longitudinal slit. Each pollen grain is surrounded by an inner intine and an outer exine. Due to incomplete nature of the exine two lateral ears are seen coming out from the pollen grain.
(b) Female i.e. Ovulate strobilus -The ovulate strobili also arise in good numbers in the axils of foliage or scale leaves borne on the dwarf shoots of female plants, and they are very much reduced. A strobilus consists of a long and slender stalk i.e. peduncle, at the tip of which usually two erect ovules i.e. megasporangia are borne only one of which usually matures into a seed while other one degenerates. Mature ovules are 1.5-2 mm in diameter. Around and partially enclosing the base of each ovule, there is an outgrowth called a “cupule” or “collar” which is probably the rudiment of the sporophyll; sometimes it may proliferate into a leaf-like structure. Sometimes 3 or 4 ovules are borne on a strobilus.
The ovule is surrounded by a single thick integument which has three layers viz. an outer fleshy layer a middle thin hard layer and an inner thin fleshy layer. A large pollen chamber is present at the apex of the nucellus. The nucellus is more or less free from the integument except at its base. The integument at the upper end forms a nucellar beak.
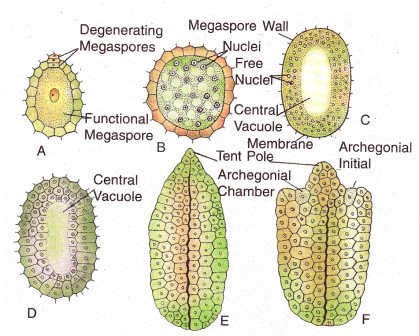
One or two megaspore mother cells become differentiated deep within the nucellus. One of the megaspore mother cells by meiosis forms a linear tetrad of megaspores, of which only the lowermost one becomes functional megaspore and other three degenerate.
Structure of the gametophytes
1. Male gametophyte- Germination of the microspore i.e. pollen grain begins while still remaining within the microsporangium. At first microspore nucleus divides to form a large antheridia initial cell and a small first prothallial cell. The larger antheridial initial cell divides again to form a persistent second prothallial cell and an antheridial cell. The first prothallial cell soon degenerates. the antheridial cell further divides to form a generative cell and a tube cell. At this 4-celled stage, pollen grains are shed from the microsporangium and they are carried to the micropyle of the ovule by wind. On reaching the micropyle they are sucked in by a drop of liquid called pollination drop and finally deposited in the pollen chamber.
The intine of the pollen grain protrudes out to form a pollen tube which acts as a haustoria. The pollen tube moves towards the female gametophyte. On its way the pollen tube absorbs the tissue of the female gametophyte. The generative cell finally divides into a stalk cell and a body cell. The body cell increases in size before its division and two blepharoplasts make their appearance at the two poles of the nucleus. Body cell next divides to form two sperm, to each of which a blepharoplast gets attached forming a spirally arranged band on which numerous flagella arise. Hence, in Ginkgo biloba sperms, are motile and Mult flagellate as in Cycas
2. Female gametophyte –The functional megaspore represents the first cell of the female gametophyte. The megaspore enlarges in size and its nucleus divides freely to form a large number of free nuclei. There is a glandular zone encircling the functional megaspore which disorganizes the nucellar tissue and makes a space for the enlarging female gametophyte. So, the megaspore is surrounded by a (i) digested, (ii) digesting and (iii) storage zones. Cell wall formation next starts at the periphery and then it proceeds, towards the centre and thereby a spongy nutritive tissue (endosperm) is formed. The endosperm is green in colour.
In addition to the megaspore membrane, the gametophyte becomes covered by another thin membrane. Two or three archegonia are formed at the upper end of the gametophyte from the archegonial initial. The archegonium consists of two-celled neck, a ventral canal cell and an egg. The archegonial clamber of Ginkgo is cylindrical surrounding a columnar mass of gametophytic tissue upon which the nucellar tissue rests like a tent of a pole-hence that columnar gametophytic tissue is called ‘tent pole’ structure.
Pollination and fertilization -These takes place as in Cycas. The archegonial chamber becomes moistened by the fluid discharged from the pollen tube. The egg and sperm nuclei fuse together to form the diploid zygote i.e. oospore.
The Embryo and Seed -After fertilization, free nuclear division of the zygote nucleus takes place giving rise to 256 free nuclei. They are evenly distributed within the cytoplasm. Next, cell walls are formed between them and the embryonic tissue is differentiated into 3 zones, such as elongated micropylar cells, larger middle cells and smaller compactly arranged basal cells.
Finally, two or sometimes three cotyledons, the radicle and the plumule having rudiments of five leaves are formed from the base. The seed normally possesses one embryo. The seed is large and is protected by a distinct three-layered seed coat (test) formed from the ovular integument. The outermost layer of the seed coat is fleshy, the middle layer is hard and stony, and the innermost turns into a thin fleshy layer. The germination of the seed is hypogeal, which finally gives rise to a new Ginkgo plant.
Economic importance:
- Ginkgo is commonly grown as an ornamental tree in the gardens all over the world. In the United States, it is also grown as a shade tree. In China and Japan, the tree is worshiped’
- The endosperm of the roasted seeds is edible. Too much of the seeds, if eaten, may prove fatal. The tree has a remarkable property of resisting fungal and bacterial attacks. Its wood is useless commercially.
- The male trees are usually preferred in the city gardens and as shade trees because the ripe seeds of female trees emit an unpleasant odour. The tree is known in Hindi as Bal Kunwari